Genetica
Disciplina che studia la trasmissione dei caratteri ereditari di generazione in generazione. Il termine fu coniato nel 1906 dal biologo britannico William Bateson. I genetisti, in particolare, studiano i meccanismi dell'ereditarietà per cui la prole generata per riproduzione sessuata assomiglia alla generazione parentale, ma non è identica a essa, e le differenze e le somiglianze ricorrono di generazione in generazione secondo uno schema ripetuto.
La nascita della genetica
La genetica è nata nel 1900, con la riscoperta degli esperimenti di
Gregor Mendel da parte di alcuni ibridatori di specie vegetali. Sebbene fosse stato pubblicato nel 1866, il fondamentale lavoro di questo monaco ceco era stato, infatti, completamente ignorato per più di quarant'anni. Mendel aveva incrociato alcune linee pure di pisello che presentavano una serie di caratteri opposti (seme liscio o rugoso, pianta alta o nana, fiore bianco o rosa ecc.) e in base ai risultati ottenuti in questi esperimenti era arrivato a formulare le regole in base alle quali questi caratteri vengono trasmessi alla generazione successiva. In particolare osservò che i fattori responsabili dei caratteri sono sempre presenti in coppia, ma solo uno dei due membri della coppia viene trasmesso alla generazione successiva; inoltre concluse che essi vengono ereditati come unità separate, ognuno indipendentemente dagli altri (vedi Leggi di Mendel). I fattori ereditari discreti ipotizzati da Mendel vennero in seguito chiamati geni.Le fasi fisiche dell'ereditarietà
 Poco
dopo
la
riscoperta
del
lavoro
di
Mendel,
alcuni
ricercatori
intuirono
che
i
meccanismi
ereditari
descritti
corrispondevano
al
comportamento
dei
cromosomi
durante
la
divisione
cellulare
e,
quindi,
suggerirono
che
le
unità
ereditarie
ipotizzate
da
Mendel,
ovverosia
i
geni,
si
trovassero
fisicamente
sui
cromosomi.
Queste
ipotesi
contribuirono
ad
accrescere
l'interesse
per
gli
studi
sulla
divisione
cellulare.
Ogni
cellula
deriva
dalla
divisione
di
una
cellula
preesistente:
ad
esempio,
tutte
le
cellule
che
costituiscono
il
corpo
di
un
essere
umano
sono
derivate
da
una
singola
cellula,
lo
zigote,
per
divisioni
successive.
Lo
zigote
si
forma
in
seguito
all'unione
dei
gameti
o
cellule
germinali
maschili
e
femminili
(uovo
e
spermatozoo)
nel
corso
della
fecondazione.
A
eccezione
di
casi
di
mutazione
o
di
altri
eventi
rari,
tutte
le
cellule
derivanti
dalla
divisione
di
un
unico
zigote
contengono
materiale
genetico
identico.
Nei
procarioti
(organismi
unicellulari
come
i batteri)
il
materiale
genetico
si
trova
libero
all'interno
della
cellula,
organizzato
in
un
unico
cromosoma
circolare.
Negli
eucarioti
(tutti
gli
altri
organismi
viventi)
il
patrimonio
ereditario
è,
invece,
racchiuso
all'interno
del
nucleo,
che
è
separato
da
una
membrana
dalle
altri
componenti
della
cellula,
localizzate
nel
citoplasma.
Nel
nucleo
il
materiale
genetico
è
organizzato
all'interno
di
più
cromosomi.
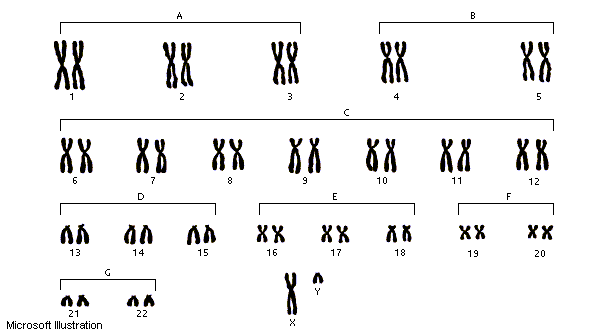
I
cromosomi
sono
strutture
filamentose,
molto
sottili,
di
forma
e
dimensioni
variabili
e
in
genere
presenti
in
coppia.
I
membri
di
ciascuna
coppia,
chiamati
cromosomi
omologhi,
sono
molto
simili
l'uno
all'altro.
A
eccezione
delle
cellule
germinali,
tutte
le
cellule
somatiche
dell'organismo
contengono
due
copie
per
ciascun
cromosoma.
Il
numero
totale
è
diverso
e
caratteristico
di
ciascuna
specie:
ad
esempio,
le
cellule
umane
contengono
23
coppie
di
cromosomi,
mentre
quelle
del
moscerino
della
frutta
(Drosophila
melanogaster)
ne
contengono
4
coppie.
Su
ogni
cromosoma
sono
localizzati
molti
geni,
la
posizione
dei
quali
su
ciascun
cromosoma
è
detta
locus.
Poco
dopo
la
riscoperta
del
lavoro
di
Mendel,
alcuni
ricercatori
intuirono
che
i
meccanismi
ereditari
descritti
corrispondevano
al
comportamento
dei
cromosomi
durante
la
divisione
cellulare
e,
quindi,
suggerirono
che
le
unità
ereditarie
ipotizzate
da
Mendel,
ovverosia
i
geni,
si
trovassero
fisicamente
sui
cromosomi.
Queste
ipotesi
contribuirono
ad
accrescere
l'interesse
per
gli
studi
sulla
divisione
cellulare.
Ogni
cellula
deriva
dalla
divisione
di
una
cellula
preesistente:
ad
esempio,
tutte
le
cellule
che
costituiscono
il
corpo
di
un
essere
umano
sono
derivate
da
una
singola
cellula,
lo
zigote,
per
divisioni
successive.
Lo
zigote
si
forma
in
seguito
all'unione
dei
gameti
o
cellule
germinali
maschili
e
femminili
(uovo
e
spermatozoo)
nel
corso
della
fecondazione.
A
eccezione
di
casi
di
mutazione
o
di
altri
eventi
rari,
tutte
le
cellule
derivanti
dalla
divisione
di
un
unico
zigote
contengono
materiale
genetico
identico.
Nei
procarioti
(organismi
unicellulari
come
i batteri)
il
materiale
genetico
si
trova
libero
all'interno
della
cellula,
organizzato
in
un
unico
cromosoma
circolare.
Negli
eucarioti
(tutti
gli
altri
organismi
viventi)
il
patrimonio
ereditario
è,
invece,
racchiuso
all'interno
del
nucleo,
che
è
separato
da
una
membrana
dalle
altri
componenti
della
cellula,
localizzate
nel
citoplasma.
Nel
nucleo
il
materiale
genetico
è
organizzato
all'interno
di
più
cromosomi.
I
cromosomi
sono
strutture
filamentose,
molto
sottili,
di
forma
e
dimensioni
variabili
e
in
genere
presenti
in
coppia.
I
membri
di
ciascuna
coppia,
chiamati
cromosomi
omologhi,
sono
molto
simili
l'uno
all'altro.
A
eccezione
delle
cellule
germinali,
tutte
le
cellule
somatiche
dell'organismo
contengono
due
copie
per
ciascun
cromosoma.
Il
numero
totale
è
diverso
e
caratteristico
di
ciascuna
specie:
ad
esempio,
le
cellule
umane
contengono
23
coppie
di
cromosomi,
mentre
quelle
del
moscerino
della
frutta
(Drosophila
melanogaster)
ne
contengono
4
coppie.
Su
ogni
cromosoma
sono
localizzati
molti
geni,
la
posizione
dei
quali
su
ciascun
cromosoma
è
detta
locus.
Il processo per il quale una cellula madre si divide in due cellule figlie identiche si chiama mitosi e comporta la duplicazione di ciascun cromosoma e la separazione delle due copie nelle due cellule figlie. Questo processo assicura che ciascuna delle due cellule figlie abbia lo stesso numero di cromosomi e, dunque, di geni della cellula madre. I processi fisiologici di crescita e sostituzione dei tessuti dell'organismo avvengono per divisione cellulare o mitosi. La riproduzione degli organismi può essere sessuata o asessuata. Quella asessuata è tipica degli organismi semplici e avviene essenzialmente per mitosi. Gli organismi superiori si riproducono, invece, per via sessuata e derivano dall'unione di due gameti, che vengono prodotti da una variante della divisione mitotica, chiamata meiosi. Questa differisce dalla mitosi per il fatto che a ciascuna delle nuove cellule viene trasmesso solo un singolo cromosoma di ciascuna coppia. In questo modo ciascun gamete viene a contenere metà dei cromosomi presenti nelle cellule somatiche. Quando avviene la fecondazione, i due gameti si uniscono a formare lo zigote e viene così restaurato il numero intero di cromosomi tipico della specie, proveniente per metà da un genitore e per metà dall'altro.
La trasmissione dei geni
L'unione dei gameti riunisce due corredi genici. Per ciascun locus genico, responsabile di un particolare carattere (ad esempio, il colore degli occhi), nel nucleo di tutte le cellule somatiche di ogni individuo sono presenti due copie, una di origine materna e una paterna (eccezioni a questa regola sono rappresentate dai cromosomi sessuali e dai caratteri legati al sesso, che vengono discussi più avanti in questo articolo). Ciascuna copia si trova nella stessa posizione, su ciascuno dei due cromosomi dello stesso tipo che generalmente stanno anche appaiati.Omozigosi ed eterozigosi
Quando le due copie del gene sono identiche, si dice che l'individuo è omozigote per quel particolare gene. Molto spesso, tuttavia, queste due copie sono diverse: ad esempio, quando un genitore ha gli occhi azzurri e l'altro ha gli occhi marroni, nella discendenza possono essere presenti due copie diverse del gene, una per ciascun colore. Se questo è il caso, si dice che l'individuo è eterozigote per quel gene. Le diverse copie di uno stesso gene sono dette alleli e occupano lo stesso locus sui cromosomi omologhi. Per ciascun gene può esistere un gran numero di alleli.
Alleli dominanti e recessivi
 Nonostante
nelle
cellule
somatiche
per
ogni
gene
siano
sempre
presenti
due
alleli,
generalmente
solo
uno
dei
due
è
manifesto
e
maschera
l'altro;
il
primo
viene
detto
dominante
e
il
secondo
recessivo.
Come
dimostrato
da
Mendel,
il
carattere
recessivo
ricompare,
nelle
generazioni
successive,
negli
individui
omozigoti
per
l'allele
considerato.
Per
convenzione
gli
alleli
sono
indicati
da
una
singola
lettera,
maiuscola
per
indicare
l'allele
dominante
(ad
esempio
A)
e
minuscola
(ad
esempio
a)
per
l'allele
recessivo.
Gli
eterozigoti
(Aa)
e
gli
omozigoti
(AA)
per
un
determinato
gene
mostrano
l'effetto
dell'allele
dominante,
mentre
gli
omozigoti
(aa)
mostrano
l'effetto
dell'allele
recessivo.
Da
quanto
descritto,
è,
quindi,
molto
importante
distinguere
tra
l'apparenza
esterna
di
un
organismo
e
i
geni
e
gli
alleli
che
esso
trasporta.
L'insieme
dei
caratteri
espressi
costituisce
il
fenotipo
dell'organismo,
mentre
il
corredo
genetico
(comprendente
anche
gli
alleli
recessivi
mascherati
dai
dominanti)
rappresenta
il
genotipo.
Ciascun
figlio
di
due
genitori
eterozigoti
(Aa)
ha
una
probabilità
su
quattro
di
essere
omozigote
(AA),
una
su
due
di
essere
eterozigote
(Aa)
e
una
su
quattro
di
essere
omozigote
(aa).
Si
noti
che
dire
che
ciascun
figlio
ha
un
quarto
di
probabilità
di
essere
omozigote
recessivo
non
equivale
a
dire
che
un
quarto
dei
figli
sarà
omozigote
recessivo.
Nonostante
nelle
cellule
somatiche
per
ogni
gene
siano
sempre
presenti
due
alleli,
generalmente
solo
uno
dei
due
è
manifesto
e
maschera
l'altro;
il
primo
viene
detto
dominante
e
il
secondo
recessivo.
Come
dimostrato
da
Mendel,
il
carattere
recessivo
ricompare,
nelle
generazioni
successive,
negli
individui
omozigoti
per
l'allele
considerato.
Per
convenzione
gli
alleli
sono
indicati
da
una
singola
lettera,
maiuscola
per
indicare
l'allele
dominante
(ad
esempio
A)
e
minuscola
(ad
esempio
a)
per
l'allele
recessivo.
Gli
eterozigoti
(Aa)
e
gli
omozigoti
(AA)
per
un
determinato
gene
mostrano
l'effetto
dell'allele
dominante,
mentre
gli
omozigoti
(aa)
mostrano
l'effetto
dell'allele
recessivo.
Da
quanto
descritto,
è,
quindi,
molto
importante
distinguere
tra
l'apparenza
esterna
di
un
organismo
e
i
geni
e
gli
alleli
che
esso
trasporta.
L'insieme
dei
caratteri
espressi
costituisce
il
fenotipo
dell'organismo,
mentre
il
corredo
genetico
(comprendente
anche
gli
alleli
recessivi
mascherati
dai
dominanti)
rappresenta
il
genotipo.
Ciascun
figlio
di
due
genitori
eterozigoti
(Aa)
ha
una
probabilità
su
quattro
di
essere
omozigote
(AA),
una
su
due
di
essere
eterozigote
(Aa)
e
una
su
quattro
di
essere
omozigote
(aa).
Si
noti
che
dire
che
ciascun
figlio
ha
un
quarto
di
probabilità
di
essere
omozigote
recessivo
non
equivale
a
dire
che
un
quarto
dei
figli
sarà
omozigote
recessivo.
Codominanza
Non è detto che vi sia sempre un allele dominante e uno recessivo. La pianta della
bella di notte, ad esempio, può avere fiori rossi, bianchi o rosa. Le piante con fiori rossi hanno due copie dell'allele R per il colore rosso dei fiori e sono, quindi, omozigoti RR. Le piante con fiori bianchi hanno due copie dell'allele r per il colore bianco dei fiori e sono, quindi, omozigoti rr. Le piante con una copia di ciascun allele, cioè gli eterozigoti Rr, hanno fenotipo rosa, intermedio tra i colori prodotti dai due alleli. Questo è un esempio del fenomeno di codominanza degli alleli.Ereditarietà dei caratteri quantitativi ed ereditarietà poligenica
Nella stragrande maggioranza dei casi i caratteri fenotipici non sono controllati da un singolo gene: solitamente un gene può influire su più di un carattere e un carattere può dipendere dall'azione di più geni. Ad esempio, per produrre il pigmento porpora nel fiore del pisello odoroso occorre l'azione di almeno due geni dominanti. Caratteri come ad esempio il peso, l'altezza, il grado di pigmentazione, che nei diversi individui presentano una gamma di variazioni quantitative continua e molto estesa, in genere dipendono da un gran numero di geni, i singoli effetti dei quali sembrano sommarsi gli uni agli altri. L'altezza di una pianta, ad esempio, potrebbe essere determinata da una serie di quattro geni: A, B, C e D. Si supponga che una pianta abbia un'altezza media di 25 cm quando il suo genotipo è aabbccdd e che ogni sostituzione con una coppia di alleli dominanti aumenti l'altezza media di circa 10 cm; in quel caso una pianta che è AABBccdd sarà alta 45 cm e una che è AABBCCDD sarà alta 65 cm. In realtà, i risultati sono raramente così regolari: geni differenti possono dare contributi differenti al valore totale e alcuni geni possono interagire in modo che il contributo di uno dipenda dalla presenza di un altro. L'eredità di caratteri quantitativi dipendenti da più geni viene detta eredità poligenica. Inoltre, quando il fenotipo è determinato, oltre che dal materiale genetico, anche da una componente ambientale, l'eredità viene detta multifattoriale.
Geni associati e crossing-over
Le seconda legge di Mendel, secondo cui i geni che controllano differenti caratteri sono ereditati indipendentemente gli uni dagli altri, è valida solo quando i geni sono portati su cromosomi diversi. Questa conclusione è dovuta agli esperimenti compiuti negli anni Trenta dal genetista americano
Thomas Hunt Morgan sul moscerino della frutta. Morgan fu in grado di dimostrare che i geni sono disposti sui cromosomi in modo lineare e che quando i geni compaiono sullo stesso cromosoma, vengono ereditati come una singola unità finché il cromosoma rimane intatto. I geni ereditati in questo modo sono detti associati. Morgan e il suo gruppo scoprirono anche che tale associazione è raramente assoluta. Le combinazioni di caratteri presenti nei genitori possono, infatti, rimescolarsi nella discendenza. Questo fenomeno è dovuto al fatto che durante la meiosi, tra le coppie di cromosomi omologhi avviene uno scambio fisico di materiale genetico, chiamato crossing-over (quando avviene, il crossing-over può essere osservato al microscopio, perché i cromosomi omologhi appaiati sono congiunti e assumono una struttura a X). Il crossing-over può avvenire con la stessa probabilità, casualmente lungo tutta la lunghezza del cromosoma. Di conseguenza la frequenza della ricombinazione tra due geni dipende dalla loro distanza sul cromosoma: se i geni sono relativamente lontani i gameti ricombinanti saranno frequenti, mentre se sono più vicini i gameti ricombinanti saranno rari. Nella discendenza derivata dall'unione di questi gameti, il crossing-over si manifesta attraverso nuove combinazioni di tratti visibili. Più crossing-over avvengono, maggiore è la percentuale di discendenti che mostra nuove combinazioni.Mappe genetiche
Dalla frequenza di ricombinazione tra due o più geni diversi, misurata in esperimenti di incrocio opportunamente progettati, è possibile dedurre la distanza che intercorre tra loro su ciascun cromosoma e costruire, così, delle mappe genetiche. L'accuratezza di queste mappe è maggiore per geni distanti che ricombinano di frequente e minore per geni vicini, che ricombinano raramente. In base ai risultati di ricerche successive agli esperimenti di Morgan, è stato dimostrato che la ricombinazione può avvenire in qualunque punto del materiale genetico, anche all'interno di un singolo gene e, grazie ad alcuni metodi messi a punto di recente, è oggi possibile individuare la ricombinazione anche tra punti molto vicini di un cromosoma.
Cromosomi sessuali e caratteri legati al sesso
 Un
altro
contributo
di
Morgan
agli
studi
genetici
fu
la
sua
osservazione,
nel
1910,
delle
differenze
sessuali
nella
trasmissione
dei
caratteri
ereditari.
Il
sesso
e
molti
dei
caratteri
a
esso
legati
sono
determinati
dall'azione
dei
geni
portati
su
una
sola
coppia
di
cromosomi.
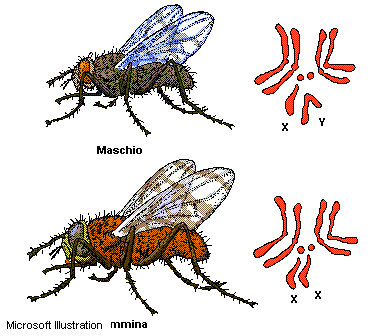
Nella
specie
umana,
ad
esempio,
vi
sono
22
coppie
di
cromosomi
uguali
in
entrambi
i
sessi
e
detti
autosomi;
la
23°
coppia,
invece,
è
quella
dei
cromosomi
sessuali,
che
nella
femmina
è
costituita
da
due
copie
del
cromosoma
X,
mentre
nel
maschio
è
formata
da
un
cromosoma
X
e
da
un
cromosoma
Y,
diversi
per
struttura
e
dimensioni.
Nella
formazione
dei
gameti,
le
cellule
uovo
femminili
contengono
sempre
un
cromosoma
X,
mentre
gli
spermatozoi
maschili
possono
contenere
un
cromosoma
X
o
un
Y.
L'unione
di
una
cellula
uovo
con
uno
spermatozoo
che
porta
un
cromosoma
X
dà
origine
a
uno
zigote
con
due
X,
cioè
a
una
femmina.
Viceversa,
dall'unione
di
una
cellula
uovo
con
uno
spermatozoo
che
porta
un
cromosoma
Y
si
ottiene
uno
zigote
XY,
cioè
un
maschio.
Su
questo
schema
fondamentale
esistono
numerose
variazioni,
caratteristiche
degli
altri
animali
e
delle
piante.
Un
altro
contributo
di
Morgan
agli
studi
genetici
fu
la
sua
osservazione,
nel
1910,
delle
differenze
sessuali
nella
trasmissione
dei
caratteri
ereditari.
Il
sesso
e
molti
dei
caratteri
a
esso
legati
sono
determinati
dall'azione
dei
geni
portati
su
una
sola
coppia
di
cromosomi.
Nella
specie
umana,
ad
esempio,
vi
sono
22
coppie
di
cromosomi
uguali
in
entrambi
i
sessi
e
detti
autosomi;
la
23°
coppia,
invece,
è
quella
dei
cromosomi
sessuali,
che
nella
femmina
è
costituita
da
due
copie
del
cromosoma
X,
mentre
nel
maschio
è
formata
da
un
cromosoma
X
e
da
un
cromosoma
Y,
diversi
per
struttura
e
dimensioni.
Nella
formazione
dei
gameti,
le
cellule
uovo
femminili
contengono
sempre
un
cromosoma
X,
mentre
gli
spermatozoi
maschili
possono
contenere
un
cromosoma
X
o
un
Y.
L'unione
di
una
cellula
uovo
con
uno
spermatozoo
che
porta
un
cromosoma
X
dà
origine
a
uno
zigote
con
due
X,
cioè
a
una
femmina.
Viceversa,
dall'unione
di
una
cellula
uovo
con
uno
spermatozoo
che
porta
un
cromosoma
Y
si
ottiene
uno
zigote
XY,
cioè
un
maschio.
Su
questo
schema
fondamentale
esistono
numerose
variazioni,
caratteristiche
degli
altri
animali
e
delle
piante.
Il cromosoma umano Y è lungo circa un terzo del cromosoma X, e, a parte il suo ruolo nel determinare il sesso maschile, non sembra essere geneticamente molto attivo. Così, molti dei geni presenti sul cromosoma X non hanno una controparte sul cromosoma Y. Sono questi i geni legati al sesso, che vengono ereditati in modo caratteristico. L'emofilia, ad esempio, è in genere causata da un gene recessivo (h) legato al sesso e portato dal cromosoma X. Una femmina con genotipo HH o Hh è generalmente sana, mentre una femmina hh ha l'emofilia. Un maschio non può mai essere eterozigote per questo gene, perché eredita una sola copia del cromosoma X e quindi un solo allele di questo gene; i genotipi possibili nel maschio sono, pertanto, H (sano) e h (malato). Quando un uomo (H) e una donna (Hh) eterozigote hanno figli, le figlie sono tutte sane, ma hanno il 50% delle probabilità di avere il genotipo Hh, come la madre, e quindi di essere portatrici del gene h. I figli maschi ereditano, invece, solo H o h e pertanto hanno il 50% di probabilità di essere malati di emofilia. Un'altra malattia ereditaria legata al sesso è il daltonismo.
L'azione dei geni
Per oltre cinquant'anni dalla nascita della genetica e dalla scoperta dei meccanismi dell'ereditarietà, molte importanti domande sull'esatta funzione dei geni negli organismi viventi sono rimaste in attesa di risposta. Ad esempio, non si conoscevano i dettagli della duplicazione dei cromosomi e della loro spartizione nelle cellule figlie durante la divisione cellulare, né si intuiva come i geni potessero determinare struttura e funzioni degli organismi viventi. I primi indizi sui meccanismi coinvolti in questi processi vennero, negli anni Quaranta, dal lavoro di due genetisti statunitensi,
George Wells Beadle e Edward Lawrie Tatum. Dai risultati di esperimenti sui funghi dei generi Neurospora e Penicillium, essi ipotizzarono che i geni contenessero al loro interno le istruzioni in codice per la formazione degli enzimi (proteine dotate di funzione catalitica nelle reazioni chimiche). Questa teoria, chiamata originariamente "ipotesi un gene-un enzima", fu in seguito ribattezzata "ipotesi un gene-un polipeptide", quando fu dimostrato che i geni contengono, in realtà, le informazioni necessarie a costruire le catene costitutive di tutte le proteine, chiamate polipeptidi. A ciascun gene corrisponde un polipeptide specifico. Questo lavoro diede l'avvio agli studi sulla natura chimica dei geni e allo sviluppo della biologia molecolare.Il DNA
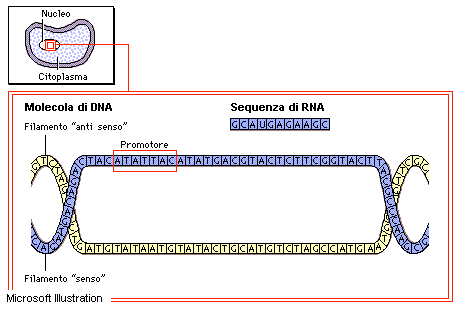
Trascrizione
La trascrizione è il processo di sintesi di un filamento di RNA messaggero, transfer o ribosomale a partire da un tratto di DNA, che viene usato come stampo. La doppia elica di quest'ultimo si srotola, rendendo accessibile uno dei due filamenti agli enzimi che controllano la reazione di sintesi. I nucleotidi necessari per costruire la nuova molecola di RNA vengono aggiunti uno per uno, in una sequenza complementare a quella dello stampo: la guanina (G) dell’RNA si lega alla citosina (C) del DNA, la citosina (C) alla guanina (G), l'adenina (A) alla timina (T) e l'uracile (U) all'adenina (A).
Negli anni Quaranta era già noto da tempo che i cromosomi sono composti da proteine e
acidi nucleici. A quel tempo, tuttavia, non tutti gli scienziati erano d'accordo su quale di questi due tipi di molecole fosse responsabile della conservazione e della trasmissione dell'informazione genetica. Nel 1944 il batteriologo canadese Oswald Theodore Avery, con un esperimento di fondamentale importanza condotto sui batteri, assegnò questo ruolo all'acido desossiribonucleico (DNA). In quegli anni si conosceva, inoltre, la composizione chimica del DNA, che era costituito da subunità, chiamate nucleotidi, a loro volta formate da un gruppo fosfato, uno zucchero (il desossiribosio) e una delle quattro basi azotate adenina (A), timina (T), guanina (G) e citosina (C). Nel 1953 il biochimico statunitense James Watson e il biofisico britannico Francis Crick, in base ai risultati di esperimenti di cristallizzazione e di diffrazione ai raggi X, elaborarono un modello tridimensionale del DNA. Secondo l'ipotesi di questi due scienziati la molecola di DNA è composta da due lunghi filamenti avvolti l'uno sull'altro come in una doppia elica o in una lunga scala a pioli. In base a questo modello i lati esterni della scala corrispondono a molecole di fosfato alternate a quelle di zucchero, mentre i pioli sono formati da coppie di basi azotate. Ciascuna base è attaccata a una molecola di zucchero presente lungo il filamento e forma un legame idrogeno con una base complementare sull'altro filamento. I legami che si possono formare sono tra l'adenina e la timina e tra la guanina e la citosina.Duplicazione del DNA
La duplicazione del DNA avviene prima di ogni divisione cellulare, in modo che le cellule figlie ricevano ciascuna una copia fedele del patrimonio genetico parentale. Per costruire una copia della molecola di DNA, i due filamenti della doppia elica si despiralizzano e si separano a livello dei legami tra le basi; a questo punto ciascun filamento funziona da stampo per l'assemblaggio di due nuovi filamenti complementari. Si formano, così, due nuove doppie eliche, ciascuna costituita da un filamento vecchio e da uno nuovo (per questo motivo la reazione di duplicazione viene detta semiconservativa). Ciascun filamento di DNA è circa 100.000 volte più lungo del cromosoma che lo contiene. Ciò è dovuto alla condensazione della molecola di DNA, che si avvolge su particelle proteiche, chiamate nucleosomi, appena visibili con i più potenti microscopi elettronici. A sua volta, la struttura formata dal DNA e dai nucleosomi si avvolge ulteriormente su se stessa più volte, fino a raggiungere lo stato di condensazione tipico del cromosoma.
Il DNA e la sintesi delle proteine
Dopo le scoperte di Beadle e Tatum e di Watson e Crick, rimaneva ancora da chiarire come il DNA potesse dirigere la costruzione delle proteine, i componenti principali della maggior parte delle strutture della cellula e le molecole fondamentali per lo svolgimento e la regolazione di quasi tutte le reazioni chimiche dell'organismo. La capacità di una proteina di essere parte di una struttura cellulare, o di agire come un enzima che catalizza una particolare reazione chimica, dipende dalla sua forma molecolare e, dunque, dalla sua composizione. Ciascuna proteina è costituita dall'unione di subunità, chiamate amminoacidi, in una o più catene polipeptidiche. Nelle cellule sono presenti venti tipi diversi di amminoacidi. Il numero, il tipo e la sequenza degli amminoacidi nella catena determinano la struttura e la funzione della proteina e sono a loro volta determinati dal gene codificante per quella specifica catena.Il codice genetico
Dal momento che si era dimostrato che le proteine sono determinate dai geni e che ciascun gene è composto da porzioni di filamenti di DNA, i ricercatori pensarono che dovesse esistere una corrispondenza tra la sequenza delle 4 basi azotate nel DNA e la sequenza dei 20 amminoacidi nelle catene polipeptidiche. In altre parole, doveva esistere un processo per trasferire l'informazione contenuta nei geni alle strutture cellulari responsabili della sintesi proteica. Siccome nel DNA compaiono solo 4 tipi di basi azotate diverse, mentre nelle proteine vi sono 20 amminoacidi, chiaramente non può valere la corrispondenza una base-un amminoacido (così le proteine potrebbero essere composte da soli 4 amminoacidi), né quella di una sequenza di due basi per amminoacido (così verrebbero specificati solo 16 amminoacidi). La sequenza minima che garantisce la determinazione di tutti gli amminoacidi presenti nelle proteine è data da combinazioni di 3 basi azotate, dette triplette o codoni, che costituiscono la base del codice genetico.
Trascrizione
L'esistenza e il ruolo biologico del codice genetico furono dimostrati dieci anni dopo la pubblicazione del modello del DNA di Watson e Crick. La specificazione di un polipeptide da parte di una molecola di DNA avviene indirettamente, attraverso l'intermediazione di una molecola nota come RNA messaggero (mRNA). L'mRNA è una replica di una porzione di DNA, la quale si despiralizza e serve da stampo per la sintesi di questa molecola. Questo processo, chiamato trascrizione, è molto simile alla duplicazione del DNA, con la differenza che l'RNA come base complementare all'adenina (A) contiene uracile (U) al posto della timina (T).
Traduzione
Mentre sta ancora avvenendo la trascrizione, l'mRNA inizia a staccarsi dal DNA. Al termine di questo processo, un'estremità del filamento della nuova molecola si inserisce, come il filo di una collana nella perla, in una struttura chiamata ribosoma. A mano a mano che il ribosoma scorre lungo l'mRNA, l'estremità del filamento si inserisce in un secondo ribosoma, poi in un terzo e così via. I ribosomi sono strutture di RNA e materiale proteico, deputate alla sintesi delle proteine. Con l'uso di microscopi elettronici ad altissima risoluzione è possibile fotografare le molecole di mRNA attaccate ai ribosomi. Un insieme di ribosomi legato a una molecola di mRNA è detto poliribosoma o polisoma. Scorrendo lungo la molecola di mRNA, il ribosoma "legge" la sequenza delle basi azotate sull'mRNA. Questo processo prende il nome di traduzione e coinvolge un terzo tipo di molecola di RNA, chiamata RNA transfer (tRNA), che da una parte porta una tripletta di nucleotidi e dall'altra un amminoacido specifico, corrispondente alla tripletta. La tripletta di ciascun tRNA aderisce alla molecola di mRNA quando vi trova una tripletta complementare. Ad esempio, la sequenza uracile-citosina-uracile (UCU) sul filamento dell'mRNA viene occupata dal tRNA contenente la tripletta adenina-guanina-adenina (AGA). In contrapposizione alla tripletta dell'mRNA, che si chiama codone, quella del tRNA prende il nome di anticodone.
Gli amminoacidi portati dal tRNA nella sequenza specificata dall'mRNA vengono, quindi, legati l'uno all'altro sui ribosomi, a formare una nuova catena polipeptidica. Una volta terminata, la catena polipeptidica si libera dal ribosoma e assume la sua forma tridimensionale specifica, determinata dalla sequenza degli amminoacidi. La forma di un polipeptide e le sue proprietà chimico-fisiche, entrambe determinate dalla sequenza amminoacidica, sono responsabili dell'eventuale unione di questa molecola ad altre catene polipeptidiche, nonché della funzione della proteina nell'organismo.
Differenze tra procarioti ed eucarioti
Nei procarioti, in cui il cromosoma è libero nel citoplasma, la traduzione può iniziare anche prima che la trascrizione sia terminata. Negli eucarioti, invece, i cromosomi sono isolati nel nucleo, mentre i ribosomi si trovano nel citoplasma, così la traduzione dell'mRNA nella proteina corrispondente può iniziare solo una volta che l'mRNA prodotto nel nucleo viene trasferito nel citoplasma. Un'altra peculiarità dei geni degli eucarioti è la presenza di sequenze di nucleotidi codificanti (esoni), interrotte da sequenze non codificanti (introni) che in alcuni casi possono essere anche cinquanta o più. Durante la trascrizione, gli introni vengono copiati insieme agli esoni su una molecola di mRNA molto grande; poi vengono eliminati da speciali enzimi nucleari e gli esoni vengono uniti l'uno all'altro in una sequenza continua, prima che l'mRNA passi nel citoplasma.
Sebbene il significato della presenza degli introni nei geni degli eucarioti non sia ancora del tutto chiaro, alcuni ricercatori ritengono che la loro esistenza permetta una serie di combinazioni di frammenti genici che andrebbe ad aumentare il numero delle possibili proteine prodotte dall'organismo. Secondo questa ipotesi, cioè, i geni degli eucarioti sarebbero costituiti da un numero relativamente basso di strutture modulari, gli esoni, in grado di combinarsi in modi diversi per dare luogo a una vastissima gamma di geni e, di conseguenza, a una grandissima varietà di strutture proteiche. Inoltre gli introni e altre sequenze non codificanti sono probabilmente coinvolti nella regolazione della quantità di polipeptidi prodotti dai geni. La scoperta degli introni fu resa possibile dai metodi di determinazione della sequenza dei nucleotidi nelle molecole di DNA e RNA, sviluppati dal biologo molecolare
Frederick Sanger. Studi sulle molecole del DNA hanno anche dimostrato la presenza, sempre negli eucarioti, di sequenze ripetute numerose volte nel materiale genetico. Alcune di queste codificano per l'RNA ribosomale, mentre altre non hanno alcuna funzione nota. Fra queste vi sono sequenze, detti trasposoni o elementi trasponibili, che sembrano in grado di saltare da una posizione all'altra di uno stesso cromosoma o dell'intero genoma.Regolazione genica
Quasi tutte le cellule di un organismo derivano per divisione cellulare da un unico zigote e contengono un identico corredo genetico. Ciononostante, le proteine sintetizzate, ad esempio, dalle cellule del
tessuto muscolare non sono necessariamente le stesse di quelle prodotte nel tessuto nervoso o in quello osseo. In altre parole, non tutti i geni del patrimonio genetico vengono espressi in tutti i tessuti dell'organismo. Quest'espressione differenziale è regolata in modo complesso da processi descritti per la prima volta da Jacques Monod e François Jacob nei batteri. Questi processi coinvolgono la presenza di sequenze regolatorie in prossimità o all'interno dei geni, le quali vengono riconosciute da specifiche molecole proteiche, con funzioni di inibizione o di attivazione della trascrizione dell'mRNA e dunque dell'espressione genica.Eredità citoplasmica
Oltre al nucleo, alcuni altri costituenti della cellula contengono DNA. Fra questi vi sono organelli citoplasmatici come i mitocondri (i produttori di energia più importanti della cellula) e i cloroplasti delle piante (responsabili della
fotosintesi clorofilliana). Questi organelli si riproducono in modo indipendente dalla cellula in cui si trovano e il loro DNA si duplica analogamente a quello nel nucleo; la maggior parte delle proteine di questi organelli è, tuttavia, codificata nel materiale genetico nucleare. Il codice genetico mitocondriale è molto simile a quello nucleare. Apparentemente queste porzioni di DNA citoplasmatico vengono più spesso ereditate dalla madre che non dal padre (nel caso di Homo sapiens esclusivamente dalla madre), poiché i gameti maschili (spermatozoi) in genere contengono meno materiale citoplasmatico di quelli femminili (cellule uovo). Altri casi di eredità apparentemente materna sono dovuti alla trasmissione di virus presenti nel citoplasma della cellula uovo.Mutazioni
Sebbene la duplicazione del DNA sia molto accurata, essa non è sempre perfetta. Raramente capitano, infatti, degli errori, per cui il nuovo frammento di DNA contiene uno o più nucleotidi diversi dall'originale. Questi errori, o mutazioni, possono avvenire in qualunque punto del DNA: se avvengono in una sequenza di DNA codificante per un particolare polipeptide, nella catena polipeptidica si può avere la variazione di un singolo amminoacido o anche un'alterazione più grave della proteina risultante. L'
anemia falcemica è, ad esempio, causata da una mutazione genetica che determina la sintesi di una molecola di emoglobina mutante, la quale differisce dalla forma normale per un singolo amminoacido. Quando una mutazione avviene nel patrimonio genetico dei gameti, essa può essere trasmessa alle generazioni successive. Il primo a parlare di mutazioni fu, nel 1901, il botanico olandese Hugo De Vries, che insieme ad altri ebbe anche il merito di riportare alla luce il lavoro di Mendel. Nel 1929 il biologo statunitense Hermann Joseph Muller osservò che i raggi X possono fare aumentare grandemente la frequenza delle mutazioni. In seguito, la lista delle sostanze mutagene si allargò ad altre forme di radiazioni, alle alte temperature e a un gran numero di composti chimici. La frequenza di mutazione aumenta, inoltre, quando alcuni geni che codificano per fattori proteici responsabili della fedeltà della duplicazione del DNA o della correzione degli errori sono a loro volta mutati. Altri fattori che possono causare mutazioni sono i trasposoni (vedi sopra).Mutazioni genetiche
La maggior parte delle mutazioni geniche è silente e non produce alcuna variazione osservabile a livello fenotipico. Raramente le mutazioni causano, invece, effetti a livello cellulare, che possono alterare in modo drammatico le funzioni generali dell'organismo. Le mutazioni non silenti compaiono generalmente in alleli recessivi e quindi i loro effetti nocivi non sono espressi se non sono presenti due alleli mutati contemporaneamente, cioè se l'individuo non è omozigote per la mutazione. Questo accade più frequentemente nei casi di inincrocio, cioè nell'accoppiamento di organismi strettamente imparentati, che possono aver ereditato lo stesso gene mutante recessivo da un comune antenato. Per questa ragione le malattie ereditarie sono più frequenti nei bambini i cui genitori sono cugini o parenti stretti, che non nella popolazione umana generale.
Mutazioni cromosomiche
La sostituzione di un nucleotide con un altro non è il solo tipo di mutazione possibile. Talvolta un nucleotide può andare perso completamente o ne può essere acquisito uno nuovo. Inoltre, possono avvenire cambiamenti più drammatici ed evidenti, come le alterazioni di forma o di numero di cromosomi. Una porzione di cromosoma può, ad esempio, staccarsi, girarsi e quindi riattaccarsi al cromosoma nello stesso punto: questa mutazione è detta inversione. Se il frammento staccato si unisce a un differente cromosoma o a una parte diversa dello stesso cromosoma, la mutazione viene chiamata traslocazione. Talvolta, un membro di una coppia di cromosomi omologhi perde un frammento di cromosoma, che viene guadagnato dall'altro membro; si dice allora che una copia ha una delezione e l'altra, una duplicazione. Le delezioni sono in genere letali negli omozigoti e spesso lo sono anche le duplicazioni. Le inversioni e le traslocazioni hanno, invece, effetti meno deleteri, sebbene possano comportare mutazioni all'interno dei geni in cui è avvenuta la rottura del cromosoma. La maggior parte di questi riarrangiamenti cromosomici è la conseguenza di errori avvenuti durante il crossing-over.
Un altro tipo di mutazione avviene quando una coppia di cromosomi omologhi non si separa alla meiosi. Questo può produrre gameti, e quindi zigoti, con cromosomi sovra- e sottonumerari. Gli individui con un cromosoma in più sono detti trisomici e quelli in cui manca un cromosoma, monosomici. Entrambe le situazioni possono dare luogo a gravi
malattie genetiche. Ad esempio, chi è affetto dalla sindrome di Down è un soggetto trisomico, cioè porta in tutte le cellule dell'organismo tre copie anziché due del cromosoma 21. Se alla meiosi non avviene la separazione dell'intero corredo cromosomico, viene prodotto un gamete con un numero di cromosomi doppio del normale. Se questo gamete si unisce a uno con un numero normale o doppio di cromosomi, la discendenza avrà rispettivamente tre o quattro set di cromosomi omologhi invece di due. Gli organismi con corredi cromosomici sovrannumerari sono detti poliploidi. La poliploidia è il solo processo conosciuto, con cui si possano originare nuove specie in una singola generazione. Poliploidi vitali e fertili si trovano quasi esclusivamente negli organismi ermafroditi, come la maggior parte delle angiosperme e alcuni animali invertebrati. Le piante poliploidi sono in genere più grandi e più resistenti dei loro normali antenati diploidi. Nell'uomo compaiono talvolta feti poliploidi, che, tuttavia, muoiono ai primi stadi di sviluppo e sono, quindi, abortiti.Genetica delle popolazioni
La genetica delle popolazioni è una disciplina, fondata da Godfrey H. Hardy e Wilhelm Weinberg, che studia come i geni sono distribuiti nelle popolazioni di organismi. In particolare essi sono responsabili della formulazione della legge di Hardy-Weinberg per determinare la frequenza di due alleli in una popolazione. I due alleli A e a vengono descritti dalle frequenze p e q, la cui somma dev'essere uguale a uno (p + q = 1). In base a questa legge, le frequenze dei tre genotipi AA, Aa e aa nella generazione successiva a quella in esame saranno rispettivamente p
2, pq2 e q2. La legge di Hardy-Weinberg vale, tuttavia, solo se la popolazione è in equilibrio genetico, cioè se sono soddisfatte le seguenti condizioni: la popolazione è isolata, cioè non è soggetta a emigrazione, né a immigrazione; l'accoppiamento tra i membri della popolazione avviene in modo casuale; le probabilità di riproduzione e sopravvivenza sono pari per tutti gli individui, cioè la popolazione non è soggetta alle forze della selezione naturale; non si verificano mutazioni; la popolazione è molto numerosa.Ereditarietà nell'uomo
Le caratteristiche fisiche dell'uomo sono per lo più influenzate sia da variabili genetiche multiple, sia da fattori ambientali. Il peso delle due componenti può essere diverso a seconda del carattere considerato. Caratteristiche come l'altezza hanno, ad esempio, una componente genetica relativamente importante, mentre altre, come il peso corporeo, sono determinate anche dalla componente ambientale. Caratteri come i
gruppi sanguigni e gli antigeni responsabili del rigetto degli organi trapiantati sono determinati esclusivamente dalla componente genetica, poiché non si conosce alcuna condizione ambientale che possa cambiare queste caratteristiche. Una componente genetica sembra, inoltre, avere un ruolo nella patogenesi di malattie come la schizofrenia, l'ipertensione e alcune forme di cancro. Gli alleli responsabili della maggior parte delle malattie genetiche rare sono recessivi. Il genoma umano contiene da 50.000 a 100.000 geni, di cui si stima che circa 4000 possono essere correlati a malattie. Nel 1990 è stato avviato il Progetto Genoma Umano, che si pone l'obiettivo di caratterizzare l'intero patrimonio genetico della specie umana. Questo gigantesco sforzo viene ripartito tra numerosi laboratori che lavorano contemporaneamente in tutte le parti del mondo ed è reso possibile dalle tecniche di ingegneria genetica e di biologia molecolare che negli ultimi anni hanno conosciuto un grande sviluppo.